13 мая 2009
Образование и выделение продуктов распада
Обмен веществ в организме заканчивается образованием продуктов распада. Они вырабатываются в клетках в результате тканевого обмена. К ним относятся углекислый газ, вода, органические вещества (например, молочная кислота), минеральные вещества — соли, железо и другие металлы.
Организм освобождается от них через органы выделения. Помимо конечных продуктов, из организма выводятся выщества, образовавшиеся при разрушении отмирающих клеток, и чужеродные соединения, попавшие вместе с пищей. Все остальные вщества, кроме газообразных, выделяются из организма в растворенном виде. Поэтому основная по весу масса выделений — вода.
Органы выделения
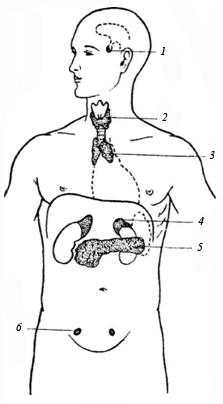
Органами выделения служат почки, кожа и легкие. Через легкие выделяются углекислый газ и пары воды. Кожа выводит из организма вещества с помощью пота и жира. Потовые железы, их около 2,5 млн., рефлекторно отделяют пот. В сутки у человека образуется около 1 л пота. Он выделяется постоянно и моментально испаряется. В состав пота входят вода, мочевина, аммиак, поваренная соль и другие вещества. Сальные железы тоже расположены в коже. Они выделяют около 20 г жира в день.
Небольшое количество веществ выделяется через кишечник. Но основную роль в выделении веществ из организма играют почки. Через них удаляются все конечные продукты обмена, кроме углекислого газа. Через почки за сутки проходит около 1000 л крови. Из нее в почках образуется моча. Она почти на 98% состоит из воды, в которой растворены мочевина и другие конечные продукты тканевого обмена, а также некоторые вещества, всасываемые из кишечника, и соли. В сутки человек вы деляет через почки 1—2 л мочи.
«Анатомия и физиология человека», М.С.Миловзорова
Читайте далее:В состав тела человека входят многие химические элементы. Содержание некоторых химических элементов в теле человека: Элементы, обязательно присутствующие в организме: Кальций Фосфор Калий Сера Хлор Натрий Магний Железо Йод Микроэлементы с незначительным содержанием в теле: Медь Марганец Цинк Фтор Кремний Мышьяк Алюминий Свинец Литий В организме они присутствуют главным образом в виде солей и некоторых кислот….
Химические превращения веществ в организме являются частью сложнейшего процесса, называемого обменом веществ. Из окружающей среды человек получав питательные вещества, воду, минеральные соли и витамины. В окружающую среду он выделяет углекислый газ, некоторое количество влаги, минеральных солей, рганических веществ. В процессе обмена веществ человек получает энергию, аккумулированную в продуктах животного и растительного происхождения, и отдает тепловую энергию…
Из общего обмена веществ 40—50% осуществляется в скелетной мускулатуре. Любая мышечная деятельность увеличивает обмен веществ в мышцах. При спокойном сидении по сравнению со спокойным лежанием он возрастает на 12%. Стояние увеличивает обмен веществ на 20%, а бег — на 400%. Причем хорошо тренированный к данному виду мышечной работы человек тратит на ее выполнении меньше энергии, чем новичок. Объясняется…
В регуляции и осуществлении обмена веществ участвуют разные отделы нервной системы. Обмен веществ и энергии, приспосабливающие его к потребностям организма, происходят под влиянием коры полушарий. Так, у тренированных спортсменов на стадионе и в спортивном зале газообмен повышается задолго до начала соревнований. Повышение обмена наблюдается и у болельщиков, несмотря на то что они только зрительно участвуют…
Выделение продуктов распада является последним этапом обмена белков, жиров и углеводов, очень важным для нормального функционирования и существования организма. Конечные и другие выделяемые продукты и некоторые вещества, введенные с лекарствами, накапливаясь в тканях, могут отравить организм. Через органы выделения они выводятся из организма. Главная функция органов выделения состоит в поддержании относительного постоянства внутренней среды организма,…
www.medkursor.ru
Обмен белков. Белки пищи под действием ферментов желудочного, поджелудочного и кишечного соков расщепляются до аминокислот, которые в тонком кишечнике всасываются в кровь, разносятся ею и становятся доступными для клеток организма. Из аминокислот в клетках разного типа синтезируются свойственные им белки. Аминокислоты, не использованные для синтеза белков организма, а также часть белков, входящих в состав клеток и тканей, подвергаются распаду с высвобождением энергии. Конечные продукты расщепления белков — вода, углекислый газ, аммиак, мочевая кислота и др. Углекислый газ выводится из организма легкими, вода — почками, легкими, кожей. Обмен углеводов. Сложные углеводы в пищеварительном тракте под действием ферментов слюны, поджелудочного и кишечного соков расщепляются до глюкозы, которая всасывается в тонком кишечнике в кровь. В печени ее избыток откладывается в виде нерастворимого в воде (как и крахмал в растительной клетке) запасного материала — гликогена. При необходимости он снова превращается в растворимую глюкозу, поступающую в кровь. Углеводы — главный источник энергии в организме. Обмен жиров. Жиры пищи под действием ферментов желудочного, поджелудочного и кишечного соков (при участии желчи) расщепляются на глицерин и ясирные кислоты (последние подвергаются омылению) . Из глицерина и жирных кислот в эпителиальных клетках ворсинок тонкого кишечника синтезируется жир, свойственный организму человека. Жир в виде эмульсии поступает в лимфу, а вместе с ней — в общий кровоток. Суточная потребность в жирах в среднем составляет 100 г. Избыточное количество жира откладывается в соединительнотканной жировой клетчатке и между внутренними органами. При необходимости эти жиры используются как источник энергии для клеток организма. При расщеплении 1 г жира выделяется наибольшее количество энергии — 38,9 кДж. Конечными продуктами распада жиров являются вода и углекисльш газ. Жиры могут синтезироваться из углеводов и белков.
Обмен веществ и энергии - это совокупность физических, химических и физиологических процессов превращения веществ и энергии в живых организмах, а также обмен веществами и энергией между организмом и окружающей средой. Обмен веществ у живых организмов заключается в поступлении из внешней среды различных веществ, в превращении и использовании их в процессах жизнедеятельности и в выделении образующихся продуктов распада в окружающую среду. Все происходящие в организме преобразования вещества и энергии объединены общим названием - метаболизм (обмен веществ) . На клеточном уровне эти преобразования осуществляются через сложные последовательности реакций, называемые путями метаболизма, и могут включать тысячи разнообразных реакций. Эти реакции протекают не хаотически, а в строго определенной последовательности и регулируются множеством генетических и химических механизмов. Метаболизм можно разделить на два взаимосвязанных, но разнонаправленных процесса: анаболизм (ассимиляция) и катаболизм (диссимиляция) . Обмен веществ начинается с поступления питательных веществ в желудочно-кишечный тракт и воздуха в легкие. Первым этапом обмена веществ являются ферментативные процессы расщепления белков, жиров и углеводов до растворимых в воде аминокислот, моно- и дисахаридов, глицерина, жирных кислот и других соединений, происходящие в различных отделах желудочно-кишечного тракта, а также всасывание этих веществ в кровь и лимфу. Вторым этапом обмена являются транспорт питательных веществ и кислорода кровью к тканям и те сложные химические превращения веществ, которые происходят в клетках. В них одновременно осуществляются расщепление питательных веществ до конечных продуктов метаболизма, синтез ферментов, гормонов, составных частей цитоплазмы. Расщепление веществ сопровождается выделением энергии, которая используется для процессов синтеза и обеспечения работы каждого органа и организма в целом. Третьим этапом является удаление конечных продуктов распада из клеток, их транспорт и выделение почками, легкими, потовыми железами и кишечником. Превращение белков, жиров, углеводов, минеральных веществ и воды происходит в тесном взаимодействии друг с другом. В метаболизме каждого из них имеются свои особенности, а физиологическое значение их различно, поэтому обмен каждого из этих веществ принято рассматривать отдельно.
Обмен белков. Белки пищи под действием ферментов желудочного, поджелудочного и кишечного соков расщепляются до аминокислот, которые в тонком кишечнике всасываются в кровь, разносятся ею и становятся доступными для клеток организма. Из аминокислот в клетках разного типа синтезируются свойственные им белки. Аминокислоты, не использованные для синтеза белков организма, а также часть белков, входящих в состав клеток и тканей, подвергаются распаду с высвобождением энергии. Конечные продукты расщепления белков — вода, углекислый газ, аммиак, мочевая кислота и др. Углекислый газ выводится из организма легкими, вода — почками, легкими, кожей. Обмен углеводов. Сложные углеводы в пищеварительном тракте под действием ферментов слюны, поджелудочного и кишечного соков расщепляются до глюкозы, которая всасывается в тонком кишечнике в кровь. В печени ее избыток откладывается в виде нерастворимого в воде (как и крахмал в растительной клетке) запасного материала — гликогена. При необходимости он снова превращается в растворимую глюкозу, поступающую в кровь. Углеводы — главный источник энергии в организме. Обмен жиров. Жиры пищи под действием ферментов желудочного, поджелудочного и кишечного соков (при участии желчи) расщепляются на глицерин и ясирные кислоты (последние подвергаются омылению) . Из глицерина и жирных кислот в эпителиальных клетках ворсинок тонкого кишечника синтезируется жир, свойственный организму человека. Жир в виде эмульсии поступает в лимфу, а вместе с ней — в общий кровоток. Суточная потребность в жирах в среднем составляет 100 г. Избыточное количество жира откладывается в соединительнотканной жировой клетчатке и между внутренними органами. При необходимости эти жиры используются как источник энергии для клеток организма. При расщеплении 1 г жира выделяется наибольшее количество энергии — 38,9 кДж. Конечными продуктами распада жиров являются вода и углекисльш газ. Жиры могут синтезироваться из углеводов и белков.
Обмен белков. Белки пищи под действием ферментов желудочного, поджелудочного и кишечного соков расщепляются до аминокислот, которые в тонком кишечнике всасываются в кровь, разносятся ею и становятся доступными для клеток организма. Из аминокислот в клетках разного типа синтезируются свойственные им белки. Аминокислоты, не использованные для синтеза белков организма, а также часть белков, входящих в состав клеток и тканей, подвергаются распаду с высвобождением энергии. Конечные продукты расщепления белков — вода, углекислый газ, аммиак, мочевая кислота и др. Углекислый газ выводится из организма легкими, вода — почками, легкими, кожей. Обмен углеводов. Сложные углеводы в пищеварительном тракте под действием ферментов слюны, поджелудочного и кишечного соков расщепляются до глюкозы, которая всасывается в тонком кишечнике в кровь. В печени ее избыток откладывается в виде нерастворимого в воде (как и крахмал в растительной клетке) запасного материала — гликогена. При необходимости он снова превращается в растворимую глюкозу, поступающую в кровь. Углеводы — главный источник энергии в организме. Обмен жиров. Жиры пищи под действием ферментов желудочного, поджелудочного и кишечного соков (при участии желчи) расщепляются на глицерин и ясирные кислоты (последние подвергаются омылению) . Из глицерина и жирных кислот в эпителиальных клетках ворсинок тонкого кишечника синтезируется жир, свойственный организму человека. Жир в виде эмульсии поступает в лимфу, а вместе с ней — в общий кровоток. Суточная потребность в жирах в среднем составляет 100 г. Избыточное количество жира откладывается в соединительнотканной жировой клетчатке и между внутренними органами. При необходимости эти жиры используются как источник энергии для клеток организма. При расщеплении 1 г жира выделяется наибольшее количество энергии — 38,9 кДж. Конечными продуктами распада жиров являются вода и углекисльш газ. Жиры могут синтезироваться из углеводов и белков.
touch.otvet.mail.ru
1. Органические вещества клетки: углеводы, жиры, белки, нуклеиновые кислоты, АТФ. Макромолекулы — крупные и сложные по строению молекулы органических соединений, состоящие из более простых молекул — «кирпичиков» . 2. Углеводы — органические соединения, состоящие из углерода, водорода и кислорода. 3. Строение углеводов. Простые углеводы — глюкоза, фруктоза. Наличие глюкозы в составе фруктов, овощей, крови человека, фруктозы — в составе фруктов и меда. Сложные углеводы — макромолекулы, состоящие из остатков молекул простых углеводов. Примеры сложных углеводов: целлюлоза (клетчатка) , крахмал, гликоген — животный крахмал, образующийся в печени. Образование молекул целлюлозы, крахмала и гликогена из остатков молекул глюкозы. Наличие в одной молекуле крахмала от нескольких сотен до нескольких тысяч остатков молекул глюкозы, а в составе молекулы целлюлозы — свыше 10000 звеньев. Прочность и нерастворимость молекул сложных углеводов. 4. Роль углеводов в организме: — запасающая — способность сложных углеводов накапливаться, образуя запас питательных веществ. Примеры: накопление крахмала в клетках клубней картофеля, корневищ многих растений; образование из молекул глюкозы и накопление в клетках печени гликогена; — энергетическая — способность молекул углеводов окисляться до углекислого газа и воды с освобождением 17,6 кДж энергии при окислении 1 г углеводов; — структурная. Углеводы — составная часть различных частей и органоидов клетки. Пример: наличие клеточной оболочки, состоящей из целлюлозы и играющей роль наружного скелета у растений. 5. Жиры — органические вещества. Гидрофоб-ность (нерастворимость в воде) — главное свойство жиров. 6. Содержание жиров в клетках в среднем от 5 до 15% , в клетках жировой ткани — до 90% . 7. Роль жиров в организме: — энергетическая — способность окисляться до углекислого газа и воды с освобождением энергии (38,9 кДж энергии при окислении 1 г жиров) ; — структурная. Жиры входят в состав плазматической мембраны; —- запасающая — способность жиров накапливаться в подкожной жировой клетчатке у животных, в семенах некоторых растений (подсолнечник, кукуруза и др.) ; — терморегуляционная: защита организма от охлаждения у ряда животных — тюленей, моржей, китов, медведей и др. ; — защитная: у ряда животных защита организма от механических повреждений, предохранение от смачивания водой перьев или волосяного покрова.
Обмен веществ и энергии - это совокупность физических, химических и физиологических процессов превращения веществ и энергии в живых организмах, а также обмен веществами и энергией между организмом и окружающей средой. Обмен веществ у живых организмов заключается в поступлении из внешней среды различных веществ, в превращении и использовании их в процессах жизнедеятельности и в выделении образующихся продуктов распада в окружающую среду. Все происходящие в организме преобразования вещества и энергии объединены общим названием - метаболизм (обмен веществ) . На клеточном уровне эти преобразования осуществляются через сложные последовательности реакций, называемые путями метаболизма, и могут включать тысячи разнообразных реакций. Эти реакции протекают не хаотически, а в строго определенной последовательности и регулируются множеством генетических и химических механизмов. Метаболизм можно разделить на два взаимосвязанных, но разнонаправленных процесса: анаболизм (ассимиляция) и катаболизм (диссимиляция) . Обмен веществ начинается с поступления питательных веществ в желудочно-кишечный тракт и воздуха в легкие. Первым этапом обмена веществ являются ферментативные процессы расщепления белков, жиров и углеводов до растворимых в воде аминокислот, моно- и дисахаридов, глицерина, жирных кислот и других соединений, происходящие в различных отделах желудочно-кишечного тракта, а также всасывание этих веществ в кровь и лимфу. Вторым этапом обмена являются транспорт питательных веществ и кислорода кровью к тканям и те сложные химические превращения веществ, которые происходят в клетках. В них одновременно осуществляются расщепление питательных веществ до конечных продуктов метаболизма, синтез ферментов, гормонов, составных частей цитоплазмы. Расщепление веществ сопровождается выделением энергии, которая используется для процессов синтеза и обеспечения работы каждого органа и организма в целом. Третьим этапом является удаление конечных продуктов распада из клеток, их транспорт и выделение почками, легкими, потовыми железами и кишечником. Превращение белков, жиров, углеводов, минеральных веществ и воды происходит в тесном взаимодействии друг с другом. В метаболизме каждого из них имеются свои особенности, а физиологическое значение их различно, поэтому обмен каждого из этих веществ принято рассматривать отдельно. Лера Котова 11 месяцев назад Знаток (275) Обмен белков. Белки пищи под действием ферментов желудочного, поджелудочного и кишечного соков расщепляются до аминокислот, которые в тонком кишечнике всасываются в кровь, разносятся ею и становятся доступными для клеток организма. Из аминокислот в клетках разного типа синтезируются свойственные им белки. Аминокислоты, не использованные для синтеза белков организма, а также часть белков, входящих в состав клеток и тканей, подвергаются распаду с высвобождением энергии. Конечные продукты расщепления белков — вода, углекислый газ, аммиак, мочевая кислота и др. Углекислый газ выводится из организма легкими, вода — почками, легкими, кожей. Обмен углеводов. Сложные углеводы в пищеварительном тракте под действием ферментов слюны, поджелудочного и кишечного соков расщепляются до глюкозы, которая всасывается в тонком кишечнике в кровь. В печени ее избыток откладывается в виде нерастворимого в воде (как и крахмал в растительной клетке) запасного материала — гликогена. При необходимости он снова превращается в растворимую глюкозу, поступающую в кровь. Углеводы — главный источник энергии в организме. Обмен жиров. Жиры пищи под действием ферментов желудочного, поджелудочного и кишечного соков (при участии желчи) расщепляются на глицерин и ясирные кислоты (последние подвергаются омылению) . Из глицерина и жирных кислот в эпителиальных клетках ворсинок тонкого кишечника синтезируется жир, свойств
touch.otvet.mail.ru
дегидрогеназа (флавиновый фермент) снова окисляет глицерол3фосфат додиоксиацетонфосфата:Глицерол3фосфат + ФАД <=> Диоксиацетонфосфат + ФАДН2.Восстановленный флавопротеин (ферментФАДН2) вводит на уровне KoQприобретенные им электроны в цепь биологического окисления и сопряженного с нимокислительного фосфорилирования, а диоксиацетонфосфат выходит из митохондрийв цитоплазму и может вновь взаимодействовать с цитоплазматическим НАДН + Н+.Таким образом, пара электронов (из одной молекулы цитоплазматического НАДН +Н+), вводимая в дыхательную цепь с помощью глицеролфосфатного челночногомеханизма, дает не 3, а 2 АТФ.Малатаспартатная челночная система для переноса восстанавливающихэквивалентов от цитозольного НАДН в митохондриальный матрикс. В дальнейшем было показано, что с помощью данного челночного механизмалишь в скелетных мышцах и мозге осуществляется перенос восстановленныхэквивалентов от цитозольного НАДН + Н+ в митохондрии.В клетках печени, почек и сердца действует более сложная малатаспартатная челночная система. Действие такого челночного механизма становитсявозможным аспартатаминотрансферазы как в цитозоле, так и в митохондриях.малатдегидрогеназы присутствиюблагодаря и
Установлено, что от цитозольного НАДН + Н+ восстановленные эквивалентысначала при участии фермента малатдегидрогеназы переносятся на цитозольныйоксалоацетат. В результате образуется малат, который с помощью системы,транспортирующей дикарбоновые кислоты, проходит через внутреннюю мембранумитохондрии в матрикс. Здесь малат окисляется в оксалоацетат, а матриксныйНАД+ восстанавливается в НАДН + Н+, который может теперь передавать своиэлектроны в цепь дыхательных ферментов, локализованную на внутренней мембранемитохондрии. В свою очередь образовавшийся оксалоацетат в присутствии глутаматаαи фермента АсАТ вступает в реакцию трансаминирования. Образующиеся аспарат и кетоглутарат с помощью специальных транспортных систем способны проходитьчерез мембрану митохондрий.Транспортирование в цитозоле регенерирует оксалоацетат, что вызывает кдействию следующий цикл. В целом процесс включает легкообратимые реакции,происходит без потребления энергии, «движущей силой» его является постоянноевосстановление НАД+ в цитозоле глицеральдегид3фосфатом, образующимся прикатаболизме глюкозы.15
znanio.ru
Присутствуют в небольших количествах в некоторых жирах (особенно в сливочном масле). Конечные продукты углеводного брожения у жвачных животных [c.152]
В отличие от углеводного обмена, где кислород служит только конечным акцептором электронов в дыхательной цепи, при окислении н-алканов кислород включается в молекулу промежуточных продуктов в самых первых реакциях. [c.238]
При нормально протекающем углеводном обмене и при преобладании распада углеводов над распадом жиров, образующаяся уксусная кислота окисляется до углекислого газа и воды. Следовательно, образование тех или иных продуктов конечного распада жирных кислот обусловливается возможностью двух путей превращения уксусной кислоты 1) ее конденсации в аце-тоуксусную кислоту и 2) ее окисления до углекислого газа и воды. [c.411]
Почти всегда при тяжелом диабете, помимо глюкозурии, наблюдается ацетон- или к е т о и у р и я, т. е. появление в моче так называемых ацетоновых (кетоновых) тел (стр. 298) ацетоуксусной кислоты, 3-0 ксимасляной кислоты и ацетона. Ацетоновые тела ( 3-оксимасляная и ацетоуксусная кислоты) рассматриваются как нормальные промежуточные продукты обмена жиров и некоторых аминокислот. В здоровом организме они окисляются до конечных продуктов обмена—СОг и НгО. Появление кетоновых тел в моче указывает на накопление их в крови в повышенных количествах ( г и п е р-к е т о н е м и я), что может произойти в результате изменения нормального те шния процессов углеводного обмена. [c.276]
Углеводный обмен во всякой живой клетке (живом веществе) представляет единый процесс одновременно протекающих связанных между собой реакций распада и синтеза органических веществ. В центре углеводного обмена у животных стоят гликогенсз и гликогенолиз, т. е. процессы образования и распада гликогена. Они протекают главным образом в печени. Гликоген может образоваться как из углеводов, так и из неуглеводных источников, таких, например, как некоторые аминокислоты, глицерин, молочная, пировиноградная и пропионовая кислоты, а также и из многих других простых соединений. Термин гликогенолиз обозначает собственно расщепление гликогена до глюкозы. Но теперь часто под этим словом понимают всю сумму процессов, ведущих к гликолитическому образованию молочной кислоты в том случае, когда исходным субстратом является не глюкоза, а гликоген. Под гликолизом понимают вообще процессы распада углеводов от начала, т. е. от глюкозы или гликогена, безразлично, и до конечных продуктов. [c.376]
Конечные продукты распада жирных кислот. В зависимости от условий, в которых протекает распад жирных кислот, конечные продукты их распада будут различными. При малом количестве распадающихся жиров и нормально протекающем углеводном обмене жирные кислоты превращаются до уксусной кислоты, которая далее окисляется до углекислого газа и воды. В случае же расстройства углеводного обмена или большого сокращения распада углеводов, например при голодании, когда сильно повышается распад жиров, конечными продуктами, кроме уксусной кислоты, являются ацетоновые тела [ 1-оксимасляная кислота, ацетоуксусная кислота и ацетон. Следует указать при этом, что прямых доказательств образования уксусной кислоты не имеется. Возможно, что процесс дальнейшего превращения образующейся уксусной кислоты при укорочении цепи жирной кислоты происходит очень быстро или образуется какое-то легко вступающее в реакцию ее производное, например ацетилфос-фат. О способности тканей животных потреблять уксусную кислоту говорит хотя бы факт отсутствия уксусной кислоты в моче при ее скармливании. [c.411]
Из азотных соединений мочи преобладает мочевина — конечный продукт обмена белков. Азот ее составляет 80—85% общего азота мочи. Количество мочевины в моче зависит от содержания протеина в кормах. При углеводных кормах мочевины в моче всегда меньше. В моче плотоядных мочевины больше, чем в моче травоядных. Б составе мочи постоянно находится креатинин. Суточное содержание его в моче связано с развитием мускулатуры. Чем она сильнее развита, тем больше выделяется с мочой креатинина. Количество креатинина в моче возрастает при напряженной мышечной работе, так как он образуется из содержащегося в мышцах креатинфосфа-та. Наряду с креатинином в моче встречается и его предшественник — креатин. В моче взрослых животных он появляется обычно после приема больших количеств мясного корма, когда его концентрация в крови превышает 1,6 мг%. В моче молодых животных креатин всегда присутствует в небольших количествах независимо от наличия мяса в кормовом рационе. При заболеваниях мышц, особенно при дистрофических процессах, содержание креатина в моче резко возрастает. [c.172]
Сравнение множества клеток самых разных типов показывает, что набор содержащихся в них ферментов во многом сходен. По-видимому, во всех живых организмах протекают в основном одни и те же метаболические процессы некоторые различия, касающиеся конечных продуктов обмена, отражают скорее наличие или отсутствие того или иного фермента, нежели изменение общего характера метаболизма. Сложные системы углеводного обмена, состоящие из ферментов, коферментов и переносчиков, образуют главный поставляющий энергию механизм у животных, растений, плесневых грибов, дрожжей и у 1большинства других микроорганизмов. [c.96]
Сравнительно недавно было показано, что карнозин также защищает мозговую ткань от образования амилоидозных отложений белка. Амилоидоз — системное заболевание, характеризующееся отложением белково-углеводных комплексов в межклеточном пространстве нервной ткани. Кроме ассоциации пептидных фрагментов в этом процессе важную роль играет сшивка пептидных фибрилл альдегидными группами редуцирующих сахаров и малоновым диальдегидом, который является вредным конечным продуктом окисления липидов. Защитный эффект карнозина при амилоидных перерождениях ткани обеспечивается, по-видимому, электрон-акцепторной активностью имидазольного кольца, препятствующей перекисному окислению. Однако эта активность проявляется только после включения гистидина в состав дипептида, имеющего значительный дипольный момент и пространственную стабилизацию кольца относительно этого диполя. Свободный гистидин такой активностью не обладает (Alberts et al., 1994). Определенную роль в дестабилизации амилоидных отложений, возможно, играет и Р-структура аланина (Iverson, 1997). [c.32]
Главный конечный продукт углеводного брожения у жвачных животных Конечный продукт углеводного брожения у жвачйых животных [c.152]
Многообразные функции печени обусловливают присутствие в ней самых разнообразных эндо- и экзогенных соединений. Это продукты белкового, углеводного и жирового обмена, биотрансформации экюгенных веществ (в том числе и токсичных), синтеза желчных кислот и т д Поэтому печень является одним из неудобных объектов для анализа и хранения. Даже после принятия всех необходимых мер, например глубокого замораживания, в конечном итоге не удается устранигь все пофешности, связанные с хранением и пробоподготовкой, при определении суперэкотоксикантов в печени. [c.203]
В жестких условиях окислительной обстановки исходное 08 подвергается глубокой трансформации. При этом обогащенный изотопом С белково-углеводный комплекс практически полностью разрушается, а ост2(вшаяся липидная часть с легким и.с.у. значительно окисляется. Вследствие действия кинетического эффекта происходит обогащение керогена изотопом С, по сравнению с продуктами окисления (в конечном счете СО ). [c.62]
Несомненно, сходные продукты образуются и при окислении других пиримидиновых нуклеозидов, но показано, что, они слишком нестабильны, чтобы их можно было выделить. Если не защищать гидроксильные группы углеводного остатка, то происходит деградация углевода с элиминирован>юм основания. Конечными азотсодержащими продуктами окисления являются гликозилмочевины, гликозилбиуреты (из цитидина) и гуанидин (и гуанина). [c.113]
Окислительное декарбоксилирование а-кетокислот в карбоновые кислоты с уменьшенной на один атом цепью углеродных атомов осуществляется при участии системы ферментов. Через ряд каталитических превращений пировиноградная кислота, являющаяся одним из продуктов углеводного обмена (в частности гликолиза), в виде продукта ее декарбоксилирования и дегидрирования — высоко макроэргического ацетил-КоА (схема 95) — вводится в цикл трикарбоновых кислот в звене превращений щавелевоуксусной кислоты в лимонную кислоту и в конечном счете окисляется в двуокись углерода и воду. Первичное расщепление пировиноградной кислоты с отделением двуокиси углерода осуществляет ТДФ. В последующих превращениях образовавшегося ацильного остатка окислительным агентом служит (+) а-липоевая кислота (ЛК, тиоктовая кислота) [376], которая сама при этом подвергается восстановительно.му ацилированию при каталитическом действии пируватдегидрогеназы в б-ацетилдигидролипоевуто кислоту. [c.421]
N-Алкил- и N-арилпиримидины, многие из которых известны, были упомянуты в разных местах предыдущих разделов. Хотя число искусственно полученных представителей этого класса соединений велико, особый интерес привлекает то обстоятельство, что большинство простейших производных пиримидина, встречающихся в природе, являются N-замещенными соединениями, причем заместителем обычно служит углеводный остаток. Так, глюкозиды, вицин и конвицин были найдены в семенах вики, а N-рибозиды или дез-оксирибозиды, полученные из урацила, цитозина и тимина, являются, как известно, продуктами гидролитического расш,епления нуклеиновых кислот подробнее эти соединения будут рассмотрены ниже. Конечно, является очевидным, что пиримидины, не имеющие в положениях 2, 4 или 6 заместителей, способных участвовать в прототропном обмене с соседними атомами азота, не могут быть замещены по N-1 или N-3 без образования четвертичных солей последние, хотя и существуют, изучены мало. При наличии в положениях 2, 4 или 6 таких заместителей как ОН или NHa алкилирование алкилгалогенидами, диметилсульфатом или диазометаном дает главным образом N-алкильные производные лактамной формы. Урацил при метилировании его диметилсульфатом образует почти с количественным выходом 1,3-диметилурацил [406], а диазометан, реагируя с тимином и 4-метилурацилом, соответственно дает [c.251]
Таким образом, функционирование гликолиза и пути Энтнера—Дудорова совместно с ЦТК, а также окислительного пентозофосфатного цикла приводит к полному окислению исходных субстратов углеводной природы. Электроны с переносчиков поступают в дыхательную цепь (см. рис. 95, В) и в зависимости от условий могут передаваться на молекулярный кислород или другие конечные акцепторы (фумарат, нитрат). Кроме того, Е. oli в анаэробных условиях в отсутствие подходящего акцептора может получать энергию, осуществляя брожение, основным продуктом которого является этанол. [c.394]
Для образования большого количества полимера требуется легкодоступный и дешевый источник углерода. Ферментация позволяет культивировать организм-продуцент в строго определенных условиях среды, контролируя, таким образом, процесс биосинтеза и влияя на тип продукта и его свойства. Специфи- чески изменяя условия роста, можно менять молекулярную массу и структуру образующегося полимера, В ряде случаев максимальная скорость синтеза полисахарида достигается в логарифмической стадии роста, в других — в поздней логарифмической или в начале стационарной. Обычно углеводными субстратами служат глюкоза и сахароза, хотя полисахариды могут образовываться и при росте микроорганизмов на н-алка-,яах( С12-61), керосине, метаноле, метане, этаноле, глицероле и этиленгликоле. Недостатком проведения процесса в ферментерах является то, что среда часто становится очень вязкой, поэтому культура быстро начинает испытывать недостаток кислорода мы все еще не умеем рассчитывать соотношение между скоростью перемешивания неньютоновских жидкостей и подачей кислорода. Необходимо также контролировать быстрые изменения pH среды. И все же упомянутый метод позволяет быстро синтезировать полимер для того, чтобы определить его физические свойства, а также дает возможность оптимизировать состав среды, главным образом в отношении эффективно- сти различных углеводных субстратов. Часто в качестве лимитирующего фактора применяют азот (соотношение углерод азот — 10 1), хотя можно использовать и другие (серу, магний, калий и фосфор). Природа лимитирующего фактора способна определять свойства полисахарида, например его вяз- костные характеристики и степень ацилирования. Так, многие оолисахариды, синтезируемые грибами, фосфорилированы. При недостатке фосфора степень фосфорилирования может уменьшаться или становиться равной нулю в этих условиях может даже измениться соотношение моносахаридов в конечном по- [c.219]
При изучении превращения двуокиси углерода (пс=1, L—0) в углеводы ( с ==6, L — 1) естественно было искать промежуточные продукты среди соединений с углеродными цепями между по—1 и 6 и обладающих уровнями восстановленности между i = 0 и 1. На подобные поиски в прошлом было затрачено много труда. В настоящее время уместно поставить следующий вопрос. Можно ли вообще рассматривать промежуточые продукты фотосинтеза как молекулы с короткими цепями, ввиду того что механизм фотосинтеза, очевидно, не включает отделения субстрата восстановления от крупной молекулы носителя в течение всего процесса восстановления Па это можно ответить, что рассуждения такого рода, конечно, не следует считать столь важными, как это казалось прежде, одпако они и не совсем бесполезны. Некоторые из химических свойств, которыми будущая углеводная молекула обладает на различных стадиях своего роста, могут быть по существу одинаковыми, независимо от того, свободна ли она, или связана с носителем. Эксперименты с радиоактивными индикаторами (стр. 251) показывают, что отделение субстрата от носителя происходит до того, как закончится его превращение в сахар. Наконец, могут существовать равновесия между свободными и связанными промежуточными продуктами, сходные с равновесиями между свободной двуокисью углерода и комплексом Og . Например, если крупная молекула органической кислоты, гидрируясь, восстанавливается вначале до альдегида, а затем до спирта, то соответственно малые молекулы — двуокись углерода, муравьиная кислота и формальдегид — могут оказаться в свободном состоянии вследствие равновесий [c.256]
Хотя сахарозу считают иногда конечным углеводным продуктом фотосинтеза, на самом деле она слулснт основным транспортируемым метаболитом во многих растениях и синтезируется в цитоплазме из ноступаюш,его туда триозофосфата. В отличие от сахарозы крахмал образуется внутри хлоропласта и встречается в большинстве листьев высших растений в качестве временного запасного продукта. Его содержание наиболее велико тогда, когда фотосинтез преобладает над потреблением ассимилированных соединений в процессе метаболизма, т. е, лист запасает крахмал днем и использует его ночью. [c.115]
chem21.info
Щавелевоуксусная кислота НООС—С(ОН)==СН —СООН. Эта кислота является продуктом нормального обмена веществ и играет существенную роль в распаде углеводов (ср. цикл лимонной кислотых, стр. 413). Как было указано выше, она образуется при окислении яблочной кислоты. Эфир ш,авелевоуксусной кислоты очень легко получается в результате сложноэфирной конденсации эфиров щавелевой и уксусной кислот, конденсирующим средством служит алкоголят натрия [c.408]
АНАЭРОБНЫЙ РАСПАД УГЛЕВОДОВ [c.204]ПРОЦЕССЫ РАСПАДА УГЛЕВОДОВ [c.363]
В результате аэробного и анаэробного распада углеводов дрожжами доставляется энергия и обеспечиваются процессы синтеза биомассы различными предшественниками. Из щавелево-уксусной и а-кетоглутаровой кислот в результате восстановительного аминирования и переаминирования образуются соответственно аспарагиновая и глутаминовая кислоты. Синтез этих двух аминокислот занимает главное место в синтезе белков из углеводов. [c.1051]
Азотное питание. Дрожжи способны синтезировать все аминокислоты, входящие в состав нх белка, непосредственно нз неорганических азотистых соединений при использовании в качестве источника углерода органических соединений — промежуточных продуктов распада углеводов, которые образуются при дыхании и брожении. [c.201]
Аэробное и анаэробное дыхание тесно связаны, и преобладание того или иного его типа зависит главным образом от наличия в среде кислорода. Общим для большинства организмов является аэробный путь распада углеводов. У зерна и клубней картофеля анаэробное дыхание усиливается при повышенных температурах и в конце периода хранения, когда активность окислительных ферментов понижена. Анаэробное дыхание может продолжаться до тех пор, пока вредные метаболиты не подавят жизнедеятельность организма. [c.43]
Магний имеет большое биологическое значение, он входит в состав хлорофилла, участвует в процессе фотосинтеза, в образовании или распаде углеводов и жиров, в превращениях фосфорных соединений. Недостаток магния в почве как микроэлемента вызывает заболевания растений (хлороз, мраморность листьев и др.). При низких содержаниях его в кормах наблюдаются заболевания и у сельскохозяйственных животных. Магниевым микроудобрением служит доломитMg Oa- [c.299]
Таким образом, для того, чтобы процесс распада органического вещества в живой клетке был наиболее энергетически выгодным, необходимо образование в ходе процесса максимально возможного количества молекул АТФ или восстановленных пиридиновых нуклеотидов. Известные нам процессы распада углеводов, несомненно, являются результатом биохимической эволюции — естественного отбора по эффективности использования энергии для жизненных процессов. Это и определяет высокую энергетическую целесообразность процессов распада углеводов. [c.364]
Ферментативный анаэробный распад углеводов исследуют при инкубации тканевого гомогената или экстракта с субстратами гликолиза (гликогеном, глюкозой, а также с промежуточными продуктами гликолиза). О процессе судят по приросту конечного продукта анаэробного превращения углеводов — лактата или убыли субстратов. Отдельные этапы изучают при добавлении в инкубационную среду ингибиторов ферментов или удалении диализом кофакторов и коферментов, необходимых для определенных реакций процесса анаэробного превращения углеводов. [c.49]
Гликолиз - один из важнейших процессов распада углеводов и образования энергии в виде макроэргических молекул АТФ. [c.78]
Гликоген — основной субстрат анаэробного распада углеводов в скелетной мускулатуре. В задаче предлагается исследовать влияние добавления АДФ на интенсивность гликогенолиза. Об интенсивности гликогенолиза судят по убыли гликогена и по образованию молочной кислоты. [c.50]
Строение и свойства углеводов, рассмотренные выше, позволяют понять их функции в живом организме. Будучи альдо- или кето-спиртами, т.е. уже частично окисленными органическими молекулами, углеводы легко вступают в реакции дальнейшего окисления. Вследствие этого основная функция углеводов - энергетическая углеводы различного строения являются основными поставщиками энергии и на их долю приходится более 50% всей вырабатываемой в организме энергии. В зависимости от того, какое вещество является акцептором электронов в ходе окисления, процессы окислительного распада углеводов делятся на анаэробные и аэробные (акцепторы электронов - различные химические субстраты в анаэробных процессах и кислород - в аэробных процессах). [c.77]
Так, хорошо известно, например, что распад углеводов в животном организме (гликолиз) протекает через сложную последовательность реакций, в которой промежуточное образование фосфорных эфиров и их превращения играют центральную роль. Далее, некоторые фосфаты сахаров входят в фер(Ментные системы. Наконец, едва ли не самыми важными природными продуктами исключительного биологического значения, представляющими собою сложные эфиры углевода, являются нуклеиновые кислоты, роль которых в биосинтезе белка и передаче наследственных признаков общеизвестна. [c.77]
В условиях аэробиоза распад углеводов до образования пировиноградной кислоты происходит так же, как и при анаэробиозе, но в отличие от него пировиноградная кислота полностью окисляется до диоксида углерода и воды в цикле трикарбоновых кислот — ЦТК (цикле Кребса, лимоннокислотном цикле). В этом цикле последовательно протекают окислительно-восстановительные реакции, в которых под действием специфических дегидрогеназ происходит перенос водорода на молекулярный кислород — конечный [c.206]
Большое значение в разнообразных процессах обмена в-в имеет ферментативное Д. Существует два типа подобных р-ций простое Д. (обратимая р-ция) и окислительное Д., в к-ром происходит сначала Д., а затем дегидрирование субстрата. По последнему типу в организме животных и растений осуществляется ферментативное Д. пировиноградной и а-кетоглутаровой к-т-промежуточных продуктов распада углеводов, жиров и белков (см. Трикарбоновых кислот цикл). Широко распространено также ферментативное Д. аминокислот у бактерий и животных. [c.19]
Почему в результате распада углеводов дрожжами доставляется энергия и обеспечиваются процессы синтеза биомассы [c.1081]
Гликолиз является эволюционно ранним процессом выработки энергии в организме в ходе распада углеводов и протекает в анаэробных условиях у микроорганизмов и при ограниченном снабжении кислородом у высших организмов. [c.78]
Переход от анаэробного пути распада углеводов к аэробному - пиру-ватдегидрогеназная реакция [c.82]
Как вырабатывается энергия в клетке в результате распада углеводов Частично АТФ, как основной макроэрг клетки, может образоваться путем субстратного фосфорилирования. Но главным процессом синтеза АТФ является окислительное фосфорилирование. [c.85]
Доказательством верности теории Митчелла является то, что существование мембранного потенциала в митохондриях стало бесспорньгм, а также то, что ионофоры (валиномицин, грамицидин, динитрофенол) создают условия для свободного перемещения ионов Н , в результате исчезает протонный градиент, и синтез АТФ прекращается. Вещества, нарушающие градиент Н , называют разобщителями окислительного фосфорилирования. Количество АТФ, синтезируемое в процессе распада углеводов Поскольку окисление одной молекулы НАДН сопровождается синтезом трех молекул АТФ, а всего в ходе гликолиза, пируватдегидрогеназной реакции и реакций ЦТК образуется десять НАДН, то всего генерируется 30 молекул АТФ, а за счет окисления двух молекул ФАДН2 образуется еще четыре молекулы АТФ, т.е. всего 34 молекулы АТФ. К этому числу следует добавить две молекулы АТФ, синтезировавшихся в гликолизе, и две молекулы ГТФ, появившихся в ЦТК за счет субстратного фосфорилирования. [c.89]
Реакции рекомбинации феноксильных радикалов приводят к образованию новых С-С- и С-О-связей в структуре лигнина, что затрудняет делигнификацию. Рекомбинация же с участием пероксильных радикалов не может дать стабильных связей. Это различие в характере взаимодействия лигнина с кислородом при избытке и недостатке последнего приводит к тому, что присутствующий в древесине и в варочном растворе кислород при обычных щелочных варках может послужить причиной конденсации фрагментов лигнина, в том числе и с участием продуктов окислительного распада углеводов. Не исключается возможность радикальной прививки лигнина к полисахаридам. Поэтому, как уже указывалось ранее, роль вводимых при делигнификации в щелочную среду химических реагентов заключается также в ингибировании окислительных процессов. [c.492]
У человека и животных на всех стадиях синтеза и распада углеводов регуляция углеводного обмена осуществляется при участии ЦНС и гормонов. [c.359]
Эти ферменты управляют процессами распада углеводов в мозговой и нервной тканях, а также при брожении различных веществ Витамин А в теле животного образуется из каротина под влиянием ферментов, т е в данном случае витамин образуется в результате ферментативной реакции [c.7]
Эти ферменты управляют процессами распада углеводов в мозговой и нервной тканях, а также при брожении различных веществ. [c.7]
С. В. Лебедев и И. А. Виноградов-Волжинский 111) сообщают о проведенных в 1911 г. опытах Л. Уббелоде и А. Воронина, Наблюдавших реакции каталитического крекинга при нагревании нефтяного масла с фуллеровой землей до температуры около 200 °С, и собственных исследованиях, в которых установили, что распад диамиленов под влиянием активированного флоридина начинается уже при 65—70 °С, заметно протекает при 90, а при 165—170 °С происходит интенсивный распад углеводо])одов. С. В. Лебедев и Г. Г. Коблянский [121 показали, что полимерные формы изобутилена под влиянием флоридина заметно распадаются уже при 130 °С. С. В. Лебедев и И. А. Лившиц [131 наблюдали распад триизобутшсена даже нри 50 °С в присутствии того же активированного флоридина. Низкотемпературный [c.154]
Открытие пути прямого окисления углеводов, или, как его называют, пентозофосфатного цикла, принадлежит О. Варбургу, Ф. Липману, Ф. Дикенсу и В.А. Энгельгарду. Расхождение путей окисления углеводов—классического (цикл трикарбоновых кислот, или цикл Кребса) и пентозофосфатного—начинается со стадии образования гексозомонофосфата. Если глюкозо-6-фосфат изомеризуется во фруктозо-6-фосфат, который фосфорилируется второй раз и превращается во фруктозо-1,6-бисфосфат, то в этом случае дальнейший распад углеводов происходит по обычному гликолитическому пути с образованием пировиноградной кислоты, которая, окисляясь до ацетил-КоА, затем сгорает в цикле Кребса. [c.353]
Очень важную роль играет степень очистки гидролизатов растительного сырья [39]. Поскольку с чистотой раствора непосредственно связана стабильность работы катализатора, а очистка является весьма дорогостоящим процессом, оптимум должен определяться по экономическому критерию. Для гидролизатов, получаемых с применением концентрированных кислот, т. е. сравнительно мало загрязненных продуктами распада углеводов, достаточной считается очистка адсорбентом (активированный уголь, коллакти-вит) и анионитами. При этом катализатор совершает в среднем 3 цикла, прежде чем выводится на регенерацию. Влияние степени очистки сырья на гидрогенолиз со стационарным катализатором пока не исследовалось, хотя для стационарного катализатора чистота сырья еще более важна, чем для суспендированного. [c.127]
Превращение белков в организме. В организмах животных и человека под влиянием ферментов (пепсина, трипси--на, эрепсина и др.) происходит гидролиз белков. В результате этого образуются аминокислоты, которые всасываются ворсинками кишечника в кровь и используются для образования белков, специфических данному организму. Синтез белков идет с поглощением энергии. Эту энергию доставляют молекулы АТФ. (Повторите из учебника Общая биология 42.) В организме одновременно с синтезом белков непрерывно происходит и полное их разрушение, вначале до аминокислот, а затем до оксида углерода (IV), аммиака, мочевины и воды. При этих процессах выделяется энергия, но Б меньшем количестве, чем при распаде углеводов и жиров. [c.21]
Напротив, окисление гексофуранозидов происходит гораздо сложнее поскольку первоначальный продукт окисления, содержащий группировку малондиальдегида, может подвергаться дальнейшему окислению. Последнее приводит в конечном счете к поглощению 6 -молей ШО4 № полному. распаду углевода с выделением спирта — агликона, четырех-молей мура.вьиной кислоты, одного моля углекислоты и одно1Го молж формальдегида (см. стр. 90). [c.89]
Основными путями распада углеводов являются гликолиз и ЦТК, но наряду с ними существуют и другие пути метаболизма углеводов. Один из них - распад глюкозо-б-фосфата до СО2 и пентоз, поэтому этот путь называют пентозофосфатным. Поскольку глюкоза является основной гексозой, распадающейся по пути гликолиза, где из нее образуется глюкозо-6-фосфат, то этот же путь получил название гексозомонофос-фатного шунта. [c.90]
О. Варбургу в 1935 г. при изучении окислительного распада углеводов впервые удалось получить в кристаллическом состоянии кофермент глюко-зо-6-фосфатдегидрогеназы. Было также установлено наличие в его составе амида никотиновой кислоты, В дальнейшем оказалось, что никотинамид является компонентом коферментов ряда ферментативных систем, участвующих во многих окислительно-восстановительных реакциях организма. Последующий период исследований ферментов ознаменовался открытием больщого числа коферментов, содержащих в своем составе те или иные витамины. Например, никотинамид — антипеллагрический витамин, входящий в состав кофермента никотинамидадениндинуклеотида [c.94]
Яблочная (гидроксибутавдиовая) кислота НООСС И(ОН)СН,СООН. Содержит один асимметрический атом углерода, поэтому возможно существование ее в виде пары энантиомеров. В природе встречается Ь-(-)-яблочная кислота (т. пл. 100 С), она содержится в ягодах н фруктах, особенно много ее в ягодах рябины и барбариса, которые используются для получения яблочной кислоты. Ь-Яблочная кислота — один из продуктов распада углеводов в живых организмах. [c.323]
Пировиноградная кислота играет важную роль в процессах обмена веществ, являясь промежуточным продуктом распада углеводов Соли и чфнры пировиноградной кислоты называются п п р у в а т а-м и. Ацетоуксусная кислота — один из продуктов биологического расщепления жиров. У больных сахарным диабетом в результате нарушения обмена вещеегв наблюдается повышенное содержание в моче ацетоуксусной кислоты н продукта ее распада — ацетона, так называемых кетоновых тел [c.332]
chem21.info
Роль углеводов в организме
В течение жизни человек съедает около 10 т углеводов. Они поступают в организм главным образом в виде крахмала. Расщепившись в пищеварительном тракте до глюкозы, углеводы всасываются в кровь и усваиваются клетками. Особенно богата углеводами растительная пища: хлеб, крупы, овощи, фрукты. Продукты животного происхождения (за исключением молока) содержат мало углеводов.
Углеводы – главный источник энергии, особенно при усиленной мышечной работе. У взрослых людей больше половины энергии организм получает за счет углеводов. Распад углеводов с освобождением энергии может идти как в бескислородных условиях, так и в присутствии кислорода. Конечные продукты обмена углеводов – углекислый газ и вода. Углеводы обладают способностью быстро распадаться и окисляться. При сильном утомлении, при больших физических нагрузках прием нескольких граммов сахара улучшает состояние организма.
В крови количество глюкозы поддерживается на относительно постоянном уровне (около 110 мг%). Уменьшение содержания глюкозы вызывает понижение температуры тела, расстройство деятельности нервной системы, утомление. Печень играет большую роль в поддержании постоянного уровня сахара в крови. Повышение количества глюкозы вызывает ее отложение в печени в виде запасного животного крахмала – гликогена, который мобилизуется печенью при снижении содержания сахара в крови. Гликоген образуется не только в печени, но и в мышцах, где его может накапливаться до 1–2 %. Запасы гликогена в печени достигают 150 г. При голодании и мышечной работе эти запасы истощаются.
Если содержание глюкозы в крови увеличивается до 0,17 %, то она начинает выводиться из организма с мочой; как правило, это происходит при употреблении с пищей большого количества углеводов. Это еще один механизм регулировки концентрации сахара в крови.
Однако в крови может наблюдаться стойкое повышение содержания сахара. Это происходит при нарушении функции желез внутренней секреции. Нарушение функционирования поджелудочной железы приводит к развитию сахарного диабета. При этом заболевании утрачивается способность тканей организма усваивать сахар, а также превращать его в гликоген и откладывать в печени. Поэтому уровень сахара в крови постоянно повышен, что приводит к усиленному выделению его с мочой.
Значение глюкозы для организма не исчерпывается ее ролью как источника энергии. Она входит в состав цитоплазмы и поэтому необходима для образования новых клеток, особенно в период роста. Входят углеводы и в состав нуклеиновых кислот.
Углеводы имеют важное значение и в обмене веществ в центральной нервной системе. При резком снижении количества сахара в крови отмечаются резкие расстройства деятельности нервной системы. Наступают судороги, бред, потеря сознания, изменение деятельности сердца. Если такому человеку ввести в кровь глюкозу или дать съесть обычный сахар, то через некоторое время эти тяжелые симптомы исчезают.
Полностью сахар из крови не исчезает даже при отсутствии его в пище, так как в организме углеводы могут образовываться из белков и жиров.
Потребность в глюкозе различных органов неодинакова. Мозг задерживает до 12 % приносимой глюкозы, кишечник – 9 %, мышцы – 7 %, почки – 5 %. Селезенка и легкие почти совсем ее не задерживают.
Обмен углеводов у детей
У детей обмен углеводов совершается с большой интенсивностью, что объясняется высоким уровнем обмена веществ в детском организме. Углеводы в детском организме служат не только основным источником энергии, но и выполняют важную пластическую роль при формировании клеточных оболочек, вещества соединительной ткани. Участвуют углеводы и в окислении кислых продуктов белкового и жирового обмена, чем способствуют поддержанию кислотно-щелочного равновесия в организме.
Интенсивный рост детского организма требует значительных количеств пластического материала – белков и жиров, поэтому образование углеводов у детей из белков и жиров ограничено. Суточная потребность в углеводах у детей высокая и составляет в грудном возрасте 10–12 г на 1 кг массы тела. В последующие годы потребное количество углеводов колеблется от 8–9 до 12–15 г на 1 кг массы. Ребенку в возрасте от 1 до 3 лет нужно давать с пищей в сутки в среднем 193 г углеводов, от 4 до 7 лет – 287 г, от 9 до 13 лет – 370 г, от 14 до 17 лет – 470 г, взрослому – 500 г.
Усваиваются углеводы детским организмом лучше, чем взрослым (у грудных детей – на 98–99 %). Вообще дети отличаются относительно большей выносливостью к повышенному содержанию сахара в крови, нежели взрослые. У взрослых глюкоза появляется в моче, если ее поступает 2,5–3 г на 1 кг массы тела, а у детей это происходит лишь при поступлении 8-12 г глюкозы на 1 кг массы тела. Прием незначительных количеств углеводов с пищей может вызвать у детей увеличение сахара в крови в два раза, но уже через 1 ч содержание сахара в крови начинает снижаться и через 2 ч полностью нормализуется.
Репликация ДНКМетоды изучения живых клеток, примеры средСпособы введения гена в клеткуЦиркодиальные ритмыМедицинская генетикаОсобенности исследования под микроскопомХарактеристика биосферыДоместикация собакиВлияние радионуклидов на организмОсобенности микобактерийПути заражения гельминтамиНейроанатомия человека
biofile.ru